MURIEL W. LAMBERT
Professor
• Ph.D.
Northwestern University, 1970
• Postdoctoral,
Harvard University, 1970
• Postdoctoral, Yale University,
1972
Research
Interests
• Cellular
responses to DNA damage
• Molecular
mechanisms of DNA repair
• Role of chromatin structure in
DNA repair
• Cancer predisposition diseases, Fanconi anemia, xeroderma
pigmentosum
|
|
Current Research
The DNA repair defect in Fanconi anemia
Damage to DNA is an important etiologic pathway for a number of important
processes such as cancer, birth defects and aging. The research programs in
my laboratory focus on mechanisms of repair of damage that occurs to cellular
DNA in mammalian cells. Of primary importance are investigations of these repair
processes in genetic diseases in which there are known DNA repair defects.
One of these diseases is the cancer prone, hematological disorder, Fanconi
anemia (FA). Patients with this disorder suffer from bone marrow failure that
leads to aplastic anemia and leukemia. These patients have a DNA repair defect,
specifically in repair of DNA interstrand cross-links. We are carrying out
research to determine the proteins involved in this repair defect and in relating
this defect to the clinical manifestations of the disorder. We have recently
shown that cells from six of the FA complementation groups tested are deficient
in the structural protein, nonerythroid a spectrin ( a SpII S *). These studies
demonstrate that a SpII S * forms a complex in the nucleus with the FANCA,
FANCC, FANCF and FANCG proteins and with XPF, a protein involved in repair
of DNA interstrand cross-links. In normal cells, these proteins co-localize
to discrete nuclear foci after the cells are damaged with a DNA interstrand
cross-linking agent. In FA cells, these nuclear foci are greatly decreased.
We have proposed a model in which a SpII S * acts as a scaffold to help recruit
and align the FA and DNA repair proteins at sites of damage and aid in their
role in the repair process. In FA cells, where there are reduced levels of
a SpII S *, there is decreased recruitment of proteins to sites of DNA damage
and decreased DNA repair.The nature of these protein interactions and their
importance in the repair process is currently being investigated. Since a spectrin
is involved in a number of different cellular processes, a deficiency in this
protein could have far reaching consequences and could thus possibly account
for some of the diverse cellular and clinical defects that have been observed
in FA cells.
Role of chromatin structure in DNA repair
Chromatin structure plays
a critical role in modulation of cellular response to DNA damage. We are
examining how it influences the ability of proteins involved in the repair
process to interact with damaged DNA. The cancer prone, DNA repair deficient
genetic disorder, xeroderma pigmentosum (XP), is being used as a model for
these studies. Cells from XP complementation group A (XPA) patients, unlike
normal human cells are defective in ability to repair damaged DNA when it
is in the form of nucleosomes. Studies in my laboratory suggest that this
is related to the mechanism by which DNA endonucleases involved in the repair
process locate sites of damage. These studies indicate that the XPA protein
may be involved in determining the mechanism of action utilized by these
endonucleases. By investigating the role that the XPA protein plays in location
of damage sites on DNA and the importance of this protein for location of target
sites on nucleosomal DNA, we expect to obtain a clearer understanding of the
mechanism of interaction of repair proteins with damaged nucleosomal DNA and
the severe consequences, such as the production of numerous cancers, that occur
in individuals when this mechanism is defective.
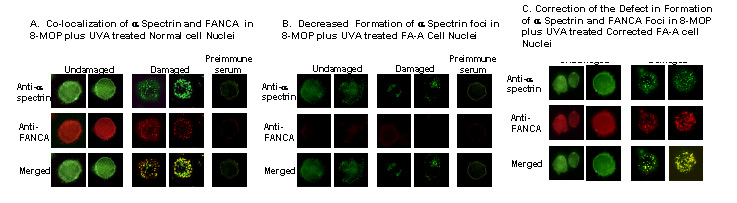
Figure 1. Localization
of a SpII S * and FANCA in the nucleus of normal and FA-A cells after treatment
with a DNA interstrand cross-linking agent, 8-MOP plus UVA light. (A) a SpII
S * and FANCA co-localize in damaged normal cell nuclei, (B) decreased formation
of a SpII S * foci and no FANCA foci in damaged FA-A cells, (C) a SpII S * and
FANCA again co-localize in nuclear foci in 8-MOP plus UVA treated corrected
FA-A cells.
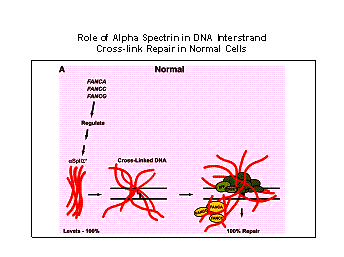
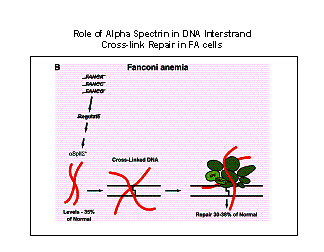
Figure 2. Model for the
role of a SpII S * in repair of DNA interstrand cross-links in (A) normal and
(B) Fanconi anemia cells. In normal cells, FANC gene products regulate the stability
of a SpII S *. a SpII S * binds to cross-linked DNA, acts as a scaffold and
aids in the recruitment and alignment of repair proteins and FANC proteins to
the site of damage thus enhancing the efficiency of the repair process. In FA
cells, defects in the FANC genes or gene products lead to decreased levels of
a SpII S * due to decreased stability of a SpII S *. Decreased levels of a SpII
S * lead to decreased binding of this protein to damaged DNA and decreased recruitment
and alignment of repair proteins at sites of damage, thus reducing the efficiency
of DNA repair in FA cells.
Representative Publications:
Lambert, M.W., and Lambert, W.C. DNA repair and chromatin structure in genetic
diseases. Prog. in Nucleic Acid Res. and Mol. Biol. 64:257-309, 1999.
Brois, D.W., McMahon, L.W., Ramos, N.I., Anglin, L.M., Walsh, C.E., and Lambert,
M.W. A deficiency in a 230 kDa DNA repair protein in Fanconi anemia complementation
group A cells is corrected by the FANCA cDNA. Carcingenesis, 20:1845-1853,
1999.
McMahon, L.W., Walsh, C.E., and Lambert, M.W. Human a spectrin II and the
Fanconi anemia proteins FANCA and FANCC interact to form a nuclear complex.
J. Biol. Chem., 274:32904-32908, 1999.
Kumaresan, K., and Lambert, M.W. Fanconi anemia, complementation group A,
cells are defective in ability to produce incisions at sites of psoralen interstrand
cross-links. Carcinogenesis, 21: 741-751, 2000.
Lambert, M.W., and Yang, L. Xeroderma pigmentosum complementation group A
protein acts as a processivity factor. Biochem. Biophys. Res. Comm., 271:782-787,
2000.
McMahon, L.W., Sangerman, J., Goodman, S.R., Kumaresan, K., and Lambert, M.W.
Human a spectrin II and the FANCA, FANCC, and FANCG proteins bind to DNA containing
psoralen interstrand cross-links. Biochemistry, 40:7025-7034, 2001.
Kumaresan, K.R., Hwang, M., Thelen, M.P., and Lambert, M.W. Contribution of
XPF functional domains to the 5' and 3' incisions produced at the site of a
psoralen interstrand cross-link. Biochemistry, 41:890-896, 2002.
Sridharan, D., Brown, M., Lambert, W.C., McMahon, L.W., and Lambert, M.W.
Nonerythroid a II spectrin is required for recruitment of FANCA and XPF to
nuclear foci induced by DNA interstrand cross-links. J. Cell Science, 116:823-835,
2003.
Lefferts, J.A., and Lambert, M.W. Fanconi anemia cell lines deficient in a
II spectrin express normal levels of a II spectrin mRNA. Biochem. Biophys.
Res. Comm., 307:510-515, 2003.
